Membrane Structure
Introduction:
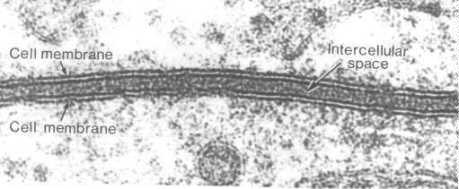
Membranes are vital because they separate the cell from the outside world. They also separate compartments inside the cell to protect important processes and events. Cellular membranes have diverse functions in the different regions and organelles of a cell. However, at the electron microscopic level, they share a common structure following routine preparative steps. The above figure shows the typical "Unit" membrane which resembles a railroad track with two dense lines separated by a clear space. This figure actually shows two adjacent plasma membranes, both of which have the "unit membrane" structure. (Above figure taken from Bloom and Fawcett, A Textbook of Histology, Chapman and Hall, N.Y., 12th edition, 1994, Figure 1-2.) To prepare for this part of the discussion on membranes, read pages 477-488 of your text (Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y. Third edition, 1994.) 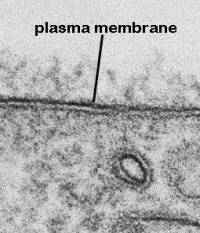 | How did early cell biologists deduce membrane structure from electron microscopic images and the knowledge that membranes were lipoprotein complexes?
Note the material projecting from the plasma membrane both inside and outside of the cell. What do you think this is? Also, outside of the cell is in the upper half of photo. Can you find a vesicle with a unit membrane structure? |
A historical perspective
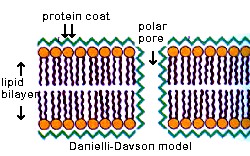
In the early 1930's-40's, Danielli and Davson studied triglyceride lipid bilayers over a water surface. They found that they arranged themselves with the polar heads facing outward. However, they always formed droplets (oil in water) and the surface tension was much higher than that of cells. However, if you added proteins, the surface tension was reduced and the membranes flattened out.
Here is a diagram of their early model of the cell membrane.
What membrane functions might be allowed by this model?
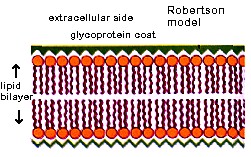
In the 1950's Robertson noted the structure of membranes seen in the above electron micrographs. He saw no spaces for pores in the electron micrographs. He hypothesized that the railroad track appearance came from the binding of osmium tetroxide to proteins and polar groups of lipids.
What is missing in Robertson's model?
In 1966, Lenard and Singer noted that over 30% of membrane proteins were twisted into an alpha helix. This made it likely that there were many spherical proteins.
Could one build a Davson-Danielli or Robertson model with spherical proteins? What would happen to the size of the membrane?
Furthermore, what would happen if you unfolded the proteins, exposing non-polar (hydrophobic amino acid side chains) groups to the aqueous environment?
What kind of energy would the cell have to expend to keep proteins flattened in this state?
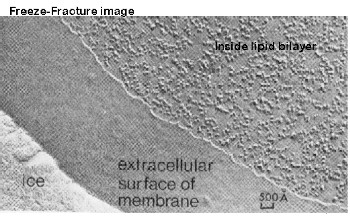
Singer studied phospholipid bilayers and found that they can form a flattened surface on water, with no requirement for a protein coat. The turning point in the modeling came with the advent of freeze fracture techniques. This figure shows the inside of a membrane and the "bumps, grooves, ridges". These were later found to be proteins.
What conclusions could you draw about membrane structure from the photo below?
|